
domingo, 29 de noviembre de 2015

APARATO TEGUMENTARIO
El sistema tegumentario está
formado por la piel y sus anexos. La piel o tegumento es el órgano que
constituye el límite del cuerpo; tapiza su superficie exterior y es la primera
barrera protectora del organismo. La piel es un claro ejemplo de cómo colaboran
entre si distintos tejido para dotar a un órgano de funciones que, de otra
forma, no sería posible cumplir.
Para realizar sus funciones
especializadas, la piel tiene requerimientos básicos que han de ser cumplidos
por los distintos tejidos. Debe ser impermeable; este carácter se lo otorga su
epitelio. Necesita resistencia mecánica, proporcionada por un entramado de
apoyo de la matriz extracelular, segregada por los fibroblastos. Necesita la
irrigación sanguínea para el transporte de sustancias; estas son intercambiadas
a través de endotelio (tejido epitelial) de los vasos sanguíneos.
Los vasos sanguíneos son
además la vía de acceso de las células inmunitarias a la piel. Las fibras
nerviosas son necesarias para transmitir información hacia el sistema nervioso
central y para distribuir señales en dirección opuesta, por ejemplo hacia las
glándulas y músculos que se ubican en la piel. Si bien la protección es la más
evidente de todas, la piel es responsable de muchas otras funciones, merced a
los tejidos que la forman.
La piel esta constituida por
tres capas: un epitelio, la epidermis, que es el tejido especializado
característico de este órgano y su parte más externa; un tejido conjuntivo, la
dermis, por debajo del anterior; y la hipodermis, su capa más profunda.
Epidermis
La epidermis es el tejido del
organismo más expuesto a las agresiones; debe estar necesariamente adaptado
para la renovación y la reparación. La epidermis es un epitelio
pluriestratificado, cuyas células reciben el nombre de queratinocitos, pues
están especializadas en la producción de queratinas, proteínas de los
filamentos intermedios. Los queratinocitos varían en las distintas capas. Los
tipos de queratinocitos son:
·
Células basales: forma la capa más interna,
adherida a la membrana basal y son las únicas que se dividen.
·
Células espinosas: se disponen en varias capas
por encima de las células basales. Deben el nombre a que sus numerosas
desmosomas, donde se anclan los filamentos de queratina, se observan como
espinas al microscopio óptico.
·
Células granulares: Sus íntimas uniones
determinan la barrera de permeabilidad que impide la pedida de fluidos. Son
además el límite entre la parte activa de la piel y la parte más superficial,
formada por células muertas.
·
Escamas: forman el estrato corneo. Se trata
de células muertas, reducidas a láminas planas, embebidas en queratina
densamente plegada.
Entre los queratinocitos de
la capa basal se ubican los melanocitos. Estas células contienen gránulos del
pigmento melanina, que le da la coloración a la piel y sus derivados. La
melanina protege de la radiación ultravioleta (RUV).
La epidermis se renueva
continuamente. Las células basales son las únicas que se reproducen. Algunas
células hijas permanecen en la capa basal, mientras que otras pasan al estrato
espinoso. Luego ascienden al estrato granuloso, donde comienzan a degradarse
convirtiéndose en escamas. Ya en el estrato corneo, las escamas se desprenden
de la superficie de la piel.
El proceso de diferenciación
está relacionado con la expresión de diferentes tipos de queratinas; a mediad
que las células pasan de una capa a otra sintetizan queratinas distintas. Las
variedades de queratina se obtienen de dos formas: porque existe una amplia
familia de genes que las codifica y por medio de un mecanismo llamado splicing
alternativo. El splicing alternativo consiste en la obtención de distintos ARNm
a partir de un mismo transcripto primario, empalmando diferentes exones. De
esta forma, un solo gen y un solo transcripto puedan dar lugar a mensajes y
proteínas diferentes.
Todo el proceso de
diferenciación de los queratinocitos dura alrededor de un mes. Las células de
la capa basal permiten la autorrenovación, pues reúnen las características de
una célula madre: no se diferencian, se dividen sin límite y cada célula hija
puede permanecer como célula madre o emprender un trayecto de diferenciación.
Una de las condiciones para que una célula basal perdure como célula madre es
que no pierda el contacto con la membrana basal.
Dermis
La dermis es el tejido
conectivo de la piel. Es un tejido laxo en su parte más superficial y denso no
modelado o irregular, en su parte más profunda. Las fibras colágenas y
elásticas de la dermis hacen a la piel resistente y deformable, al mismo
tiempo. La dermis forma elevaciones llamadas papilas que se proyectan hacia la
dermis. En algunas partes del cuerpo, como las palmas de las manos y plantas de
los pies, la epidermis acompaña los pliegues de la dermis, formando surcos y
elevaciones: son las huellas que se utilizan para identificar a las personas.
La dermis recibe abundante
irrigación sanguínea; como el epitelio es tejido no vascularizados,
los nutrientes y desechos que deben llegar a la epidermis o que deben
abandonarla, difunden a través de la matriz de la dermis. El diámetro de los
vasos sanguíneos de la piel es controlado por el sistema nervioso autónomo. La
vasodilatación y la vasoconstricción se utilizan para aumentar o disminuir el
flujo sanguíneo de la piel. La vasodilatación permite la disipación del calor
corporal hacia el ambiente. Por lo contrario, cuando es necesario conservar el
calor, o bien derivar un mayor caudal sanguíneo hacia otros órganos, se produce
la vasoconstricción.
Hipodermis
También llamada panículo
adiposo o tejido celular subcutáneo, la hipodermis es la capa más profunda de
la piel. Está formada por tejido adiposo, especializado en la reserva de
grasas. El grosor de la hipodermis varía según las zonas del cuerpo y el estado
nutricional del individuo. La grasa de la hipodermis es la reserva energética a
largo plazo. La capa de grasa subcutánea actúa como aislante térmico, ayudando
a conservar el calor corporal.

Glándulas
En la piel hay tres tipos de
glándulas que se desarrollan a partir de la epidermis: las sebáceas, las
sudoríparas y las mamarias. Las gandulas, de la naturaleza epitelial, se
desarrollan como invaginaciones de la epidermis. Su cuerpo queda ubicado en el
espesor de la dermis, pero se mantienen conectadas con la superficie a través
de sus conductos.
Las glándulas sebáceas
secretan sebo, una materia grasa cuya misión es impermeabilizar y lubricar la
piel y os pelos. Además, los ácidos grasos presentes en el sebo impiden el
crecimiento de algunos microbios.
Las sebáceas pertenecen al
tipo de glándulas denominada holocrinas (holo: total, crino: secreción)
aquellas que pierden la totalidad del contenido celular cuando liberan su
secreción. Sus conductos desembocan en el folículo piloso.
Las glándulas sudoríparas
secretan sudor, que se libera por los poros de la piel. Como el sudor contiene
desechos, pero también sustancias que cumplen un papel de defensa contra los
microbios, puede decirse que es al mismo tiempo una excreción y una secreción. Además
tiene una función refrigerante.
Las glándulas sudoríparas
pertenecen a dos tipos: merocrinas, cuando su secreción es liberada sin que se
vea afectada la integridad celular, y apocrinas, si liberan parte del
citoplasma apical junto con su secreción. Las glándulas mamarias son glándulas
apocrinas.

Faneras
Las faneras son los anexos
de la piel, como pelos o uñas. Son de origen epidérmico, y sus diferentes consistencias
dependen del tipo de queratinas sintetizadas. Los pelos se originan de una
invaginación epidérmica, el folículo piloso, en cuya parte más profunda se
ubican las células madre que se reproducen continuamente haciendo crecer el
pelo. La parte del pelo que emerge está formada por células muertas. El color
del pelo depende de los depósitos de melanina, de la cantidad de burbujas de
aire que contiene y de su textura.
Los pelos se erizan cuando
se contrae el musculo piloerector, que se extiende desde las papilas hasta el
folículo. La contracción es involuntaria y aparece como respuesta al frio o al
temor. Las uñas se originan y crecen de modo similar a los pelos. En la piel se
localizan diferentes tipos de terminaciones nerviosas sensoriales
especializadas en la recepción de estímulos táctiles, térmicos y dolorosos.
INFOGRAFIA

APARATO REPRODUCTOR MASCULINO
Órganos genitales externos
-Testículos
- escroto
-
pene
Órganos genitales internos
-
conductos deferentes
- vesículas seminales
-
Conductos eyaculadores
-
próstata
Glándulas genitales auxiliares
-
Glándulas bulbo retrales
TESTÍCULOS
Los testículos son dos glándulas ovoides, una
a cada lado del pene, de unos 5 cm de largo y 2,5 cm de diámetro y con un peso
de 10-15 gramos. Producen las células germinales masculinas o espermatozoides y
las hormonas sexuales masculinas o andrógenos. Constituyen las gónadas
masculinas y tienen el mismo origen embriológico que los ovarios o gónadas
femeninas. En el interior de cada testículo, los espermatozoides se forman en
varios cientos de túbulos seminíferos que se unen para formar una red de
canales que recibe el nombre de rete testis.
Los túbulos seminíferos
contienen dos tipos de células, las células espermatogénicas, que darán lugar a
los espermatozoides y las células de Sertoli encargadas del mantenimiento del
proceso de formación de espermatozoides o espermatogénesis. En el tejido
conjuntivo situado en los espacios que separan a los túbulos seminíferos
adyacentes hay grupos de células llamadas células de Leydig que secretan
testosterona, el andrógeno más importante.
EPIDIDIMOS
Los epidídimos son dos
estructuras en forma de coma de unos 4 cm de longitud, cada una de las cuales
se encuentra adosada a las superficies superior y posterior-lateral de cada
testículo. Cada epidídimo tiene un cuerpo que consiste en el conducto del
epidídimo que está muy contorneado y en donde los espermatozoides son
almacenados para pasar las etapas finales de su maduración, y una cola o cola
del epidídimo que se continúa con el conducto deferente que transporta el
esperma hacia el conducto eyaculador para su expulsión hacia la uretra.
ESCROTO
El escroto es un saco
cutáneo exterior que contiene los testículos y está situado
posteroinferiormente en relación al pene e inferiormente en relación a la sínfisis
del pubis. Consta de:
·
piel: rugosa, de color oscuro
·
fascia superficial o lámina de tejido
conjuntivo que contiene una hoja de músculo liso con el nombre de músculo
dartos cuyas fibras musculares están unidas a la piel y cuya contracción
produce arrugas en la piel del escroto.
La localización exterior del
escroto y la contracción de sus fibras musculares regulan la temperatura de los
testículos porque la producción normal de espermatozoides requiere una
temperatura inferior en unos 2-3 grados a la temperatura corporal.
Cuerpo
del pene: es la parte pendular libre, cubierta por piel muy fina,
de color oscuro y poco adherida. Está compuesto por tres cuerpos cilíndricos de
tejido cavernoso eréctil, encerrados en una cápsula fibrosa. Dos de los cuerpos
eréctiles que son los cuerpos cavernosos, están situados a ambos lados del
pene, en la parte posterior del órgano. El otro cuerpo eréctil que es el cuerpo
esponjoso, se encuentra anteriormente en el pene, en el plano medio, contiene
la uretra esponjosa y la mantiene abierta durante la eyaculación.
Raíz
del pene: es la parte superior, de sujeción del pene y contiene
los dos pilares que se insertan a cada lado del arco púbico. Cada pilar del
pene está rodeado por el músculo isquiocavernoso. El bulbo del pene es la
porción ensanchada de la base del cuerpo esponjoso por donde penetra la uretra
esponjosa en el cuerpo esponjoso, y está unido a los músculos profundos del
periné. La contracción de todos estos músculos esqueléticos permite la
eyaculación.
En la punta del pene, el
cuerpo esponjoso forma el glande que cubre los extremos libres de los cuerpos
cavernosos. Cerca del extremo final del glande se encuentra el orificio de la
uretra esponjosa u orificio externo de la uretra. La piel y las fascias del
pene se prolongan como una doble capa de piel dando lugar al prepucio, que
cubre el glande en una extensión variable.
El tejido eréctil de los
cuerpos cavernosos y del cuerpo esponjoso consiste en unos espacios venosos
entrelazados e intercomunicados entre sí. Cuando se produce excitación sexual
las fibras nerviosas parasimpáticas, que provienen de la médula espinal sacra,
estimulan la producción y liberación de óxido nítrico (NO) que provoca la
relajación del músculo liso de las arteriolas que proporcionan sangre a estos
espacios venosos y como consecuencia la sangre fluye y los llena, de modo que
los cuerpos cavernosos se agrandan y se vuelven rígidos, y el pene se pone en
erección.
La eyaculación es la
liberación brusca de semen desde la uretra al exterior debido a un reflejo
simpático coordinado por la región lumbar de la médula espinal. Como parte de
ese reflejo, el esfínter de músculo liso situado en la base de la vejiga
urinaria se cierra, evitando que la orina sea expulsada durante la eyaculación
y que el semen ingrese en la vejiga urinaria.
El semen es una mezcla de
espermatozoides y líquido seminal, el cual está formado por las secreciones de
los túbulos seminíferos, las vesículas seminales, la próstata y las glándulas
bulbouretrales. Es decir, es una suspensión de espermatozoides en los líquidos segregados
por dichas estructuras. El volumen de semen de una eyaculación normal es de
unos 2,5-5 ml y contiene unos 50-150 millones de espermatozoides por cada ml.
Los conductos deferentes son
2 tubos musculares de pared gruesa que comienzan en la cola del epidídimo de
cada lado y terminan en el conducto eyaculador. Transportan esperma desde el
epidídimo al conducto eyaculador de su lado.
Las vesículas seminales son
2 largos tubos de unos 15 cm. de longitud que están enrollados y forman unas
estructuras ovaladas en la base de la vejiga, por delante del recto. Sería más
apropiado llamarlas glándulas seminales ya que producen una secreción espesa y
alcalina que contiene fructosa, prostaglandinas y diversas proteínas, que se
mezcla con el esperma a medida que éste pasa a lo largo de los conductos
eyaculadores. El conducto de cada vesícula seminal se une con el conducto
deferente de su lado para formar el conducto eyaculador.
Cada uno de los dos
conductos eyaculadores es un tubo delgado, mide de 2 a 2,5 cm. de longitud y se
forma cerca del cuello de la vejiga por la unión del conducto de la vesícula
seminal y el conducto deferente de su lado. Ambos conductos eyaculadores viajan
juntos a medida que pasan a través de la próstata y van a desembocar en la
uretra prostática en donde expulsan el semen inmediatamente antes de que sea
expulsado al exterior desde la uretra.
La próstata es la mayor
glándula accesoria del sistema reproductor masculino con un tamaño similar al
de una pelota de golf. Se sitúa en la pelvis por debajo de la vejiga urinaria y
detrás de la sínfisis del pubis y rodea la primera porción de la uretra que,
por ello, se llama uretra prostática. El líquido prostático es lechoso y
levemente ácido y contiene ácido cítrico, enzimas proteolíticos y sustancias
antibióticas.
Las glándulas bulbouretrales
son 2 y tienen el tamaño de un guisante, también reciben el nombre de glándulas
de Cowper. Se encuentran debajo de la próstata, póstero - laterales en relación
a la uretra membranosa. Sus conductos (2 - 3 cm.) se abren en la porción
superior de la uretra esponjosa. Durante la excitación sexual secretan un
líquido alcalino al interior de la uretra que protege a los espermatozoides,
neutralizando la acidez de la uretra y moco que lubrifica el extremo del pene y
las paredes de la uretra, disminuyendo el número de espermatozoides que
resultan dañados por la eyaculación.
La uretra masculina es un
tubo muscular que transporta la orina y el semen hasta el orificio externo de
la uretra o meato uretral, localizado en el extremo del glande. Con propósitos
descriptivos, la uretra masculina se divide en 3 partes:
·
Uretra prostática
·
Uretra membranosa
·
Uretra esponjosa
La uretra prostática tiene
unos 3 cm de longitud y comienza en el orificio interno de la uretra, en el
trígono de la vejiga. Desciende a través de la glándula prostática y en su
pared posterior desembocan los conductos de la glándula prostática y los
conductos eyaculadores. La uretra membranosa es la porción más corta de la
uretra con 1 cm de longitud aproximadamente. Está rodeada por el esfínter
uretral y a cada lado se encuentra una glándula bulbouretral. La uretra
esponjosa es la porción más larga de la uretra con unos 15 cm de longitud y
atraviesa toda la longitud del pene. Termina en el orificio externo de la
uretra que comunica con el exterior y es la porción más estrecha y menos
distensible de la uretra con un diámetro aproximado de unos 5 mm. En la parte
superior de esta uretra desembocan los conductos de las glándulas
bulbouretrales.
La espermatogénesis o formación
de los espermatozoides, tiene lugar en los túbulos seminíferos de los
testículos en donde se encuentran las células germinales en diversas fases de
desarrollo. Las células germinales son células indiferenciadas llamadas
espermatogonias, Cada espermatogonia aumenta de tamaño y se convierte en un
espermatocito primario, Al dividirse el espermatocito primario da lugar a dos
espermatocitos secundarios, De cada espermatocito secundario se originan dos
células hijas llamadas espermátides, Por último, se produce la transformación
de cada una de las espermátides en un espermatozoide.
El espermatozoide humano
maduro es una célula alargada (de unas 60 micras de largo) y delgada y consiste
en una cabeza y una cola. En la cabeza se encuentra el núcleo, el acrosoma, que
contiene numerosos enzimas que ayudan al espermatozoide a penetrar en el
ovocito secundario y así conseguir la fecundación. La cola es móvil con una
gran cantidad de mitocondrias en la parte proximal, y la parte restante es, en
realidad, un largo flagelo que contiene microtúbulos con una estructura similar
a la de los cilios, que sirven para que el espermatozoide pueda avanzar.
También es un órgano
endocrino cuyas células de Leydig producen y secretan la hormona testosterona
que es un andrógeno y la principal hormona masculina. Las
funciones de la testosterona son:
·
La testosterona interviene en el desarrollo
embrionario del aparato genital externo masculino
·
La testosterona mantiene la espermatogénesis,
actuando sobre receptores situados en las células de Sertoli, Controla la
velocidad y el mantenimiento de la espermatogénesis actuando sobre las células
de Sertoli, pero solamente cuando sobre éstas ya ha tenido lugar la acción de
la hormona folículoestimulante (FSH) de la adenohipófisis.
·
La testosterona es responsable de diversas
características del sexo masculino, como algunos aspectos del comportamiento,
mayor masa muscular, modificaciones de la laringe. También del desarrollo de
las glándulas accesorias del tracto reproductor masculino. Asimismo contribuyen
a la líbido o impulso sexual.

INFOGRAFIA

APARATO REPRODUCTOR FEMENINO
ANATOMÍA DEL APARATO REPRODUCTOR FEMENINO.

Los órganos genitales de la mujer están más protegidos que el de los hombres por estar internamente colocados. El ovario es el encargado de producir un óvulo cada mes. El óvulo va desde la trompa de Falopio hasta el útero. Mensualmente, un óvulo sale del cuerpo a través de la menstruación. Sin embargo, si el óvulo es fecundado, éste se implanta en la cavidad uterina donde se formará un feto.
la menstruacion
La mujer sangra cada mes debido a un óvulo que sale del cuerpo. Las adolescentes comienzan la menstruación entre los 12 a 14 años. La ausencia de éste puede indicar la presencia de un tumor, por ello debe ser examinada por desordenes reproductivos. Una semana antes de la menstruación, la mujer puede sufir del síndrome premenstrual, en el cual ella se pone agresiva y le dan dolores de cabeza y de ovarios.
La menopausia
Es la desaparición de las hormonas femeninas. Erróneamente se cree que con la menopausia, la mujer sufre de traúmaticos cambios emocionales y sexuales. Aunque hay un cambio físico al cual la mujer debe de acostumbrarse, la vida sexual muchas veces es más activa ya que no hay preocupación de medidas preventivas. Los trastornos emocionales en la mayoria de las veces provienen de la idea de vejez. Un tratamiento hormonol puede aplicar, sin embargo se debe de tener cuidado ya que los efectos secundarios pueden producir depresiones nerviosas y hemorrágeas. Para prevenir la osteoporosis, se recomienda un tratamiento con estrógenos.
Está formado por los órganos genitales internos, los órganos genitales externos y las mamas.
Órganos genitales internos

En donde distinguimos, entre otras, las siguientes estructuras:
|

INFOGRAFIA

SISTEMA URINARIO
También se le conoce con el nombre de Aparato Excretor. Está formado por una serie de estructuras cuya función principal es recoger y eliminar todas las sustancias de desecho resultantes de las reacciones bioquímicas que tienen lugar en el organismo.
Los órganos principales de este aparato son los riñones que forman la orina a partir de un proceso de filtración de la sangre. Por tanto, las funciones del aparato urinario se pueden resumir como:
- Formación de la orina en el riñón. La formación y eliminación de la orina contribuye a la regulación del medio interno.
- El riñón también se comporta como una glándula endocrina secretando una hormona, la eritropoyetina, que es necesaria en la producción de glóbulos rojos (hematopoyesis). También produce renina que participa en la regulación de la presión arterial.
- Transporte de la orina hasta la vejiga urinaria a través de los uréteres.
- Almacenamiento de la orina en la vejiga.
- Eliminación de la orina a través de la uretra.
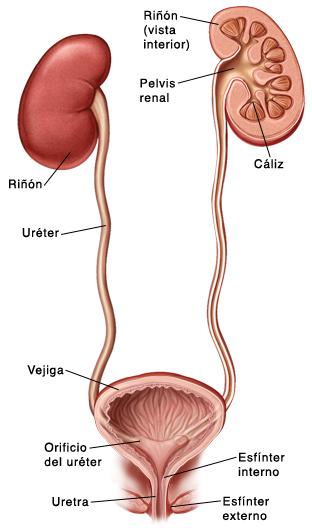
Anatomía del aparato urinario
Constituido por dos riñones, dos uréteres, la vejiga y la uretra.
Riñones
Son dos órganos macizos, uno derecho y otro izquierdo, situados en la región lumbar, uno a cada lado de la columna vertebral y algo por delante de ésta. Su tamaño es de 11 x 3 x 5 cm, aproximadamente y su peso oscila entre 110 y 180 gramos. En forma de habichuela el riñón presenta dos bordes, uno externo y otro interno en el que se localiza una hendidura central denominada hilio renal.
El riñón derecho está ligeramente más bajo que el izquierdo, ya que el hígado lo desplaza hacia abajo

Si realizamos un corte en un riñón en sentido vertical, se observarán las siguientes partes:
- Corteza renal: Es la porción más externa del mismo. De aspecto uniforme. Tiene aproximadamente 1 cm de espesor y rodea la médula.
- Médula renal: Es la porción más interna del riñón. Tiene aspecto estriado y está formada por pirámides cónicas denominadas Pirámides de Malphigio. El número de pirámides oscila entre 8 y 18 en cada riñón. La base de cada pirámide está orientada hacia el exterior y el vértice hacia el hilio renal. En el vértice de la misma se localiza la papila renal.
El hilio renal es una hendidura situada en el borde interno del riñón. A través del hilio renal penetran en el rinón la arteria renal y nervios y salen la vena renal y uréter.La zona de la corteza renal situada entre cada dos pirámides se denomina columna de Bertin.Un lóbulo renal está formado por la pirámide renal y la correspondiente zona de corteza que la rodea.Las pirámides renales se unen por su extremo convexo en los llamados cálices menores, que son de 8 a 10 por pirámide, y que a su vez se unen para formar de 2 a 3 cálices mayores. Los cálices mayores se unen entre sí para formar la pelvis renal. La pelvis renal desemboca en el uréter.La unidad estructural y funcional del riñón se denomina Nefrona. En cada riñón hay entre 1 y 3 millones de nefronas. Cada nefrona está formada por:
Corpúsculo renal: Constituido por el Glomérulo y la Cápsula de Bowman. El glomérulo está formado a su vez por una tupida red de capilares sanguíneos envueltos por una membrana denominada Cápsula de Bowman. En el interior de esa cápsula entra una arteriola, denominada arteriola aferente y sale otra llamada arteriola eferente. La Cápsula de Bowman es una membrana de doble hoja, que se invagina sobre sí misma para alojar al glomérulo, creando en su interior un espacio, el espacio de Bowman, donde se recoge la orina filtrada del glomérulo.
Túbulo Contorneado Proximal (TCP): Es la continuación del corpúsculo renal. Presenta dos zonas, una situada en la corteza renal, que presenta muchas sinuosidades alrededor del corpúsculo renal, y otra situada en la zona medular del riñón, mucho más recta que la primera. La pared del TCP está formada por una capa de células epiteliales apoyadas sobre una membrana basal.
Asa de Henle: En forma de U. Está formada por una porción descendente y delgada y una porción ascendente que en la primera parte del trayecto es delgada mientras que en la segunda es gruesa.
Túbulo Contorneado Distal (TCD): Es la continuación del Asa de Henle.
Túbulo colector (TC): Es un tubo recto. Se reúnen entre sí para desaguar en los cálices de la pelvis renal.
La cápsula de Bowman, TCP y TCD están situados en la corteza renal. Asa de Henle y TC se sitúan en la médula renal.
.jpg)
Hay nefronas que ocupan en el riñón una posición cortical mientras otras se sitúan en posición yuxtamedular.
Uréteres
Son dos largos tubos, uno izquierdo y otro derecho, que comunican por su extremo superior con la pelvis renal y por su extremo inferior con la vejiga urinaria. Tienen una longitud aproximada de 30 cm.
La pared ureteral está formada por las siguientes capas: una capa mucosa, que tapiza internamente la luz del tubo, una capa de músculo liso y una capa externa o adventicia.
Vejiga
Es una especie de saco membranoso que actúa como reservorio de orina entre cada dos micciones. Situada detrás de la sínfisis del pubis tiene forma de pera. Presenta una base ancha de forma triangular, el trígono de lietaud, en cuyos vértices superiores desembocan los uréteres. En el vértice inferior tiene su comienzo la uretra.
Uretra
Representa la parte final de las vías urinarias. En la mujer la uretra es muy corta (4 cm aproximadamente). En el varón mide unos 20 cm aproximadamente.
En el varón hay que diferenciar tres segmentos, a saber: uretra prostática, uretra membranosa y uretra cavernosa.
- la uretra prostática mide unos 3 cm de longitud,. Atraviesa el espesor de la próstata y en ella desemboca la próstata y los dos conductos deferentes.
- la uretra membranosa es muy corta (2,5 cm), y presenta un engrosamiento de fibras musculares esqueléticas que corresponde al esfínter externo. Dicho esfínter está controlado voluntariamente.
- la uretra cavernosa discurre en el espesor del músculo del mismo nombre, mide unos 15 cm y termina en el meato urinario.
La unión de la uretra con la vejiga presenta un engrosamiento muscular denominado esfínter uretral interno, formado por fíbras musculares dispuestas en haces espirales, circulares y longitudinales que constituyen el músculo detrusor de la vejiga.
Formación de la orina
La homeostasis consiste en el mantenimiento constante del medio interno, sin modificación de los parámetros bioquímicos. Esta función se lleva a cabo gracias al riñón que se comporta como una estación depuradora de la sangre que atraviesa los glomérulos renales.
La formación de la orina definitiva que produce el aparato excretor es el resultado de tres mecanismos diferentes, a saber:
- Filtración glomerular.
- Reabsorción tubular.
- Secreción tubular.
Filtración glomerular
La sangre que atraviesa los glomérulos es sometida a un proceso de filtración, que la hace pasar desde la luz de los capilares glomerulares hacia la luz de la cápsula de Bowman. La barrera de filtración la forman: endotelio de los capilares glomerulares, la membrana basal y la capa de células epiteliales (podocitos) de la cápsula de Bowman. No todos los componentes de la sangre son capaces de atravesar esta barrera. Así, en condiciones normales, las células sanguíneas y las moléculas de medio y alto peso molecular no son filtradas.
El filtrado glomerular está compuesto fundamentalmente por agua, electrolitos y moléculas de distinta naturaleza pero de bajo peso molecular, manteniendo una concentración similar a la del plasma sanguíneo. En el filtrado glomerular apenas existen proteínas, dado que su elevado peso molecular dificulta que atraviesen la barrera glomerular.
Esta filtración se produce debido a la presión efectiva de filtración, que es la fuerza neta que permite el paso de agua y solutos a través de la barrera de filtración.
La Presión efectiva de filtración es el resultado de:
- La diferencia de presiones entre la presión hidrostática de la luz de los capilares glomerulares y la presión hidrostática de la luz de la cápsula de Bowman. Esta presión favorece la salida de los líquidos hacia la cápsula de Bowman.
- La Presión oncótica del capilar glomerular. Se debe a las proteínas del plasma que por su carga eléctrica de superficie son capaces de atraer y retener agua y solutos dentro de la luz capilar. Esta presión evita en cierto grado la salida de un mayor volumen de líquidos hacia la cápsula de Bowman.
Los riñones humanos filtran al día aproximadamente 180 litros y sin embargo se eliminan en condiciones normales 1,5 I de orina.
Reabsorción tubular
En condiciones normales el riñón reabsorbe el 99% del agua y del sodio filtrados.
También reabsorbe moléculas importantes que son aprovechadas en el metabolismo general y que por su bajo peso molecular son filtradas, tal como ocurre con la glucosa, aa, etc.
a) TCPLa reabsorción se realiza con el paso de líquidos desde la luz tubular al espacio intersticial inmediato y de ahí a la sangre (capilares).En este fragmento se reabsorbe entre el 65-70% del sodio filtrado. La reabsorción de sodio se acompaña de la reabsorción de cloro y bicarbonato para mantener la neutralidad eléctrica.Se reabsorbe la totalidad de la glucosa y aa filtrados.Se reabsorbe el 50% del potasio filtrado bien por un mecanismo activo o pasivo con predominio del segundo mecanismo.Como consecuencia de la reabsorción de los anteriores, se produce una disminución de la osmolaridad del líquido filtrado y aumenta la del líquido reabsorbido al espacio intersticial. Se crea así una diferencia de concentración entre ambos compartimentos que favorece la reabsorción pasiva del agua.También se reabsorbe el 50% de la urea filtrada por un mecanismo pasivo.b) Asa de HenleEn la rama descendente del Asa se reabsorbe agua y también se produce la secreción neta de urea, que pasa del espacio intersticial a la luz del Asa para ser eliminada por la orina. El líquido que queda en la luz del Asa se vuelve hipertónico al perder el agua y mantener los solutos.En la porción delgada de la rama ascendente del Asa de Henle se reabsorbe sodio y cloro de forma pasiva, debido al gradiente de concentración, que es mayor en el Asa que en el espacio intersticial. Al final de este fragmento el líquido es isotónico.En la porción gruesa de la rama ascendente del Asa se reabsorbe por transporte activo cloro al que acompañan sodio y potasio. El líquido tubular se vuelve hipotónico.c) TCDHay un intercambio de sodio por potasio. El sodio es reabsorbido y el potasio secretado desde el espacio intersticial a la luz del TCD. Este mecanismo de intercambio está controlado por la acción de la hormona llamada aldosterona. El paso del sodio al espacio intersticial se acompaña del paso de cloro. El líquido resultante sigue siendo hipotónico.d) TCEn este segmento tiene lugar la regulación definitiva del agua a favor de gradiente. El epitelio del túbulo es impermeable al agua y para que lo sea necesita de la acción hormonal.Se reabsorbe agua hacia el espacio interticial por mecanismo activo que es controlado por la hormona antidiurética (ADH). El líquido del túbulo colector se vuelve hipertónico.Se reabsorbe entre el 60-70% de la urea.
Aclaramiento renal
El riñón actúa como una estación depuradora retirando de la sangre gran cantidad de metabolitos y sustancias tóxicas producidos en el metabolismo general del organismo.
El aclaramiento en él permite valorar esta capacidad renal. Mide la capacidad de los riñones para eliminar una sustancia del plasma. Se puede definir como el volumen de plasma que por la acción depuradora de los riñones queda libre de esa sustancia en la unidad de tiempo.
En condiciones normales el proceso de formación de la orina en sus tres fases de filtración, reabsorción y secreción ayuda a mantener el equilibrio hidroelectrolítico y ácido base de la sangre. El aclaramiento renal permite valorar el buen funcionamiento del riñón.

INFOGRAFIA

SISTEMA ENDOCRINO
Las hormonas son sustancias químicas producidas por el cuerpo que controlan numerosas funciones corporales.
Las hormonas actúan como "mensajeros" para coordinar las funciones de varias partes del cuerpo. La mayoría de las hormonas son proteínas que consisten de cadenas de aminoácidos. Algunas hormonas son esteroides, sustancias grasas producidas a base de colesterol.
Las hormonas van a todos lugares del cuerpo por medio del torrente sanguíneo hasta llegar a su lugar indicado, logrando cambios como aceleración del metabolismo, aceleración del ritmo cardíaco, producción de leche, desarrollo de órganos sexuales y otros.
El sistema hormonal se relaciona principalmente con diversas acciones metabólicas del cuerpo humano y controla la intensidad de funciones químicas en las células. Algunos efectos hormonales se producen en segundos, otros requieren varios días para iniciarse y durante semanas, meses, incluso años.

Funciones que controlan las hormonas
Entre las funciones que controlan las hormonas se incluyen:
- Las actividades de órganos completos.
- El crecimiento y desarrollo.
- Reproducción
- Las características sexuales.
- El uso y almacenamiento de energía
- Los niveles en la sangre de líquidos, sal y azúcar.
Metabolismo Hormonal
El hígado y los riñones desempeñan un papel fundamental en la depuración y excreción de estas hormonas, pero poco se sabe acerca del proceso detallado de su metabolismo. La vida media de la prolactina es de 12 minutos; la de la LH y FSH es cercana a la hora, mientras que la HCG tiene una vida media de varias horas. Si el contenido de ácido siálico es mayor, más prolongada es la supervivencia de la hormona en la circulación.
Fábrica de hormonas
Las encargadas de producir las hormonas son las glándulas endocrinas. Dentro de ellas, el primer lugar lo ocupa sin duda la hipófisis o glándula pituitaria, que es un pequeño órgano de secreción interna localizado en la base del cerebro, junto al hipotálamo. Tiene forma ovoide (de huevo) y mide poco más de diez milímetros. A pesar de ser tan pequeñísima, su función es fundamental para el cuerpo humano, por cuanto tiene el control de la secreción de casi todas las glándulas endocrinas.
La hipófisis está formada por dos glándulas separadas, conocidas como adenohipófisis y neurohipófisis. La primera corresponde al lóbulo anterior y la segunda al lóbulo posterior. Se comunica anatómica y funcionalmente a través de la sangre con el hipotálamo, lo que articula una gran coordinación entre el sistema nervioso y el endocrino.
La relación hipotálamo-hipófisis es bastante particular, puesto que, a diferencia del resto del sistema nervioso, en que las neuronas se relacionan directamente con su efector (órgano terminal que distribuye los impulsos nerviosos que recibe, activando la secreción de una glándula o contracción de un músculo), en la hipófisis las neuronas hipotalámicas no hacen contacto directo con sus efectoras. Estas últimas pasan a la sangre y alcanzan la adenohipófisis a través de una red capilar que se extiende entre el hipotálamo y la hipófisis anterior. En consecuencia, los núcleos hipotalámicos son fundamentales para el normal funcionamiento de la hipófisis.
Regulación de las hormonas
La regulación de hormonas en general incluye tres partes importantes:
El hígado y los riñones desempeñan un papel fundamental en la depuración y excreción de estas hormonas, pero poco se sabe acerca del proceso detallado de su metabolismo. La vida media de la prolactina es de 12 minutos; la de la LH y FSH es cercana a la hora, mientras que la HCG tiene una vida media de varias horas. Si el contenido de ácido siálico es mayor, más prolongada es la supervivencia de la hormona en la circulación.
Fábrica de hormonas
Las encargadas de producir las hormonas son las glándulas endocrinas. Dentro de ellas, el primer lugar lo ocupa sin duda la hipófisis o glándula pituitaria, que es un pequeño órgano de secreción interna localizado en la base del cerebro, junto al hipotálamo. Tiene forma ovoide (de huevo) y mide poco más de diez milímetros. A pesar de ser tan pequeñísima, su función es fundamental para el cuerpo humano, por cuanto tiene el control de la secreción de casi todas las glándulas endocrinas.
La hipófisis está formada por dos glándulas separadas, conocidas como adenohipófisis y neurohipófisis. La primera corresponde al lóbulo anterior y la segunda al lóbulo posterior. Se comunica anatómica y funcionalmente a través de la sangre con el hipotálamo, lo que articula una gran coordinación entre el sistema nervioso y el endocrino.
La relación hipotálamo-hipófisis es bastante particular, puesto que, a diferencia del resto del sistema nervioso, en que las neuronas se relacionan directamente con su efector (órgano terminal que distribuye los impulsos nerviosos que recibe, activando la secreción de una glándula o contracción de un músculo), en la hipófisis las neuronas hipotalámicas no hacen contacto directo con sus efectoras. Estas últimas pasan a la sangre y alcanzan la adenohipófisis a través de una red capilar que se extiende entre el hipotálamo y la hipófisis anterior. En consecuencia, los núcleos hipotalámicos son fundamentales para el normal funcionamiento de la hipófisis.
Regulación de las hormonas
La regulación de hormonas en general incluye tres partes importantes:
- heterogeneidad de la hormona
- regulación hacia arriba y hacia abajo de los receptores
- regulación de la adenil-ciclasa.
Los factores de crecimiento son producidos por expresión local de genes. Operan por unión a receptores en la membrana celular. Los receptores generalmente contienen un componente intracelular con tirosina-quinasa. Otros factores actúan a través de segundos mensajeros, tales como el AMPc y el fosfoinositol.
Los factores de crecimiento requieren condiciones especiales para actuar; para inducir la mitogénesis se requiere la exposición secuencial a varios de ellos, con limitantes importantes en cantidad y tiempo de exposición. Pueden actuar en forma sinérgica con hormonas; por ejemplo el IGF-I en presencia de FSH induce receptores para LH.
Regulación de arriba hacia abajo
"La modulación positiva o negativa de los receptores por hormonas homólogas es conocida como regulación hacia arriba y hacia abajo"
Poco se conoce sobre la regulación hacia arriba, pero se sabe que hormonas como la prolactina y la GnRH pueden aumentar la concentración de sus propios receptores en la membrana.
La principal forma biológica como las hormonas peptídicas controlan el número de receptores y por ende, la actividad biológica, es a través del proceso de internalización. Esto explica el por qué de la secreción pulsátil de las gonadotropinas para evitar la regulación hacia abajo.
"Cuando hay concentraciones elevadas de hormona en la circulación, el complejo hormona-receptor se mueve hacia una región especial en la membrana, el hueco revestido (coated pit)". A medida que esta región se va llenando sufre el proceso de endocitosis mediada por receptores. Esta región de la membrana celular es una vesícula lipídica que está sostenida por una canasta de proteínas específicas llamadas clatrinas.
Cuando está completamente ocupada la vesícula es invaginada, se separa e ingresa a la célula como una vesícula cubierta, llamada también receptosoma. Es transportada a los lisosomas donde sufre el proceso de degradación. El receptor liberado puede ser reciclado y reinsertado en la membrana celular; a su vez, tanto el receptor como la hormona pueden ser degradados disminuyendo la actividad biológica.
Este proceso de internalización no solo es utilizado para el control de la actividad biológica sino para transporte intracelular de sustancias como hierro y vitaminas.
Los receptores de membrana han sido divididos en dos clases. Los de clase I son utilizados para modificar el comportamiento celular por regulación hacia abajo; son ocupados por FSH, LH, HCG, GnRH, TSH, TRH e insulina. Los receptores de clase II son utilizados para ingreso de sustancias indispensables para la célula y para remover noxas; por ejemplo son usados por la LDL para el transporte de colesterol a las células esteroidogénicas.
Heterogeneidad
Las glicoproteínas tales como FSH y LH no son proteínas únicas sino una familia de formas heterogéneas (isoformas) con diversa actividad biológica e inmunológica. Las isoformas tienen variación en la vida media y peso molecular.
Esta familia de glicopéptidos incluye la FSH, LH, TSH y HCG. Todas son dímeros compuestos de dos subunidades polipeptídicas glicosiladas, las subunidades a y b. Todas comparten la subunidad a que es idéntica, conformada por 92 aminoácidos. Las cadenas b difieren tanto en los aminoácidos como en el contenido de carbohidratos, lo cual les confiere especificidad.
El factor limitante en la producción hormonal está dado por la disponibilidad de cadenas b, ya que las a se encuentran en cantidad suficiente a nivel tisular y sanguíneo.
Las glicoproteínas pueden variar en su contenido de carbohidratos. La remoción de residuos de la FSH lleva a la producción de compuestos capaces de unirse al receptor pero no de desencadenar acciones biológicas.
La prolactina consta de 197 a 199 aminoácidos; tiene también variaciones estructurales que incluyen glicosilación, fosforilación y cambios en unión y carga eléctrica. Se encuentran varios tamaños que han llevado a utilizar términos como pequeña, grande y gran-gran prolactina.
Todas estas modificaciones e isoformas llevan a que el inmunoanálisis no siempre pueda reflejar la situación biológica.
están compuestos principalmente por proteínas, pero tienen modificaciones secundarias de carbohidratos y pueden estar selectivamente inmersos en la membrana lipídica, también pueden estar fosforilados, o formar oligómeros por puentes de disulfuro o interacciones covalentes.
Para ejercer su acción, todas las hormonas deben unirse a su receptor específico, estas uniones inician mecanismos intracelulares que conllevan las respuestas celulares. Las hormonas esteroideas y tiroideas son liposolubles y entran a las células libremente y se unen a las proteínas del citosol. Los complejos resultantes translocan al núcleo donde se unen a elementos regulatorios en el DNA estimulando o inhibiendo la transcripción de genes específicos. Todas las demás hormonas se unen a los receptores celulares localizados en la membrana de las células diana. Esta unión disipara uno o más de las vías de transducción que llevan a las respuestas celulares.
Clases y clasificación de Hormonas
Inicialmente las hormonas se clasificaban en tres grupos de acuerdo a su estructura química: hormonas peptídicas y proteicas, las hormonas asteroideas y las hormonas relacionadas con aminoácidos.En vertebrados se clasifican en:
Los factores de crecimiento requieren condiciones especiales para actuar; para inducir la mitogénesis se requiere la exposición secuencial a varios de ellos, con limitantes importantes en cantidad y tiempo de exposición. Pueden actuar en forma sinérgica con hormonas; por ejemplo el IGF-I en presencia de FSH induce receptores para LH.
Regulación de arriba hacia abajo
"La modulación positiva o negativa de los receptores por hormonas homólogas es conocida como regulación hacia arriba y hacia abajo"
Poco se conoce sobre la regulación hacia arriba, pero se sabe que hormonas como la prolactina y la GnRH pueden aumentar la concentración de sus propios receptores en la membrana.
La principal forma biológica como las hormonas peptídicas controlan el número de receptores y por ende, la actividad biológica, es a través del proceso de internalización. Esto explica el por qué de la secreción pulsátil de las gonadotropinas para evitar la regulación hacia abajo.
"Cuando hay concentraciones elevadas de hormona en la circulación, el complejo hormona-receptor se mueve hacia una región especial en la membrana, el hueco revestido (coated pit)". A medida que esta región se va llenando sufre el proceso de endocitosis mediada por receptores. Esta región de la membrana celular es una vesícula lipídica que está sostenida por una canasta de proteínas específicas llamadas clatrinas.
Cuando está completamente ocupada la vesícula es invaginada, se separa e ingresa a la célula como una vesícula cubierta, llamada también receptosoma. Es transportada a los lisosomas donde sufre el proceso de degradación. El receptor liberado puede ser reciclado y reinsertado en la membrana celular; a su vez, tanto el receptor como la hormona pueden ser degradados disminuyendo la actividad biológica.
Este proceso de internalización no solo es utilizado para el control de la actividad biológica sino para transporte intracelular de sustancias como hierro y vitaminas.
Los receptores de membrana han sido divididos en dos clases. Los de clase I son utilizados para modificar el comportamiento celular por regulación hacia abajo; son ocupados por FSH, LH, HCG, GnRH, TSH, TRH e insulina. Los receptores de clase II son utilizados para ingreso de sustancias indispensables para la célula y para remover noxas; por ejemplo son usados por la LDL para el transporte de colesterol a las células esteroidogénicas.
Heterogeneidad
Las glicoproteínas tales como FSH y LH no son proteínas únicas sino una familia de formas heterogéneas (isoformas) con diversa actividad biológica e inmunológica. Las isoformas tienen variación en la vida media y peso molecular.
Esta familia de glicopéptidos incluye la FSH, LH, TSH y HCG. Todas son dímeros compuestos de dos subunidades polipeptídicas glicosiladas, las subunidades a y b. Todas comparten la subunidad a que es idéntica, conformada por 92 aminoácidos. Las cadenas b difieren tanto en los aminoácidos como en el contenido de carbohidratos, lo cual les confiere especificidad.
El factor limitante en la producción hormonal está dado por la disponibilidad de cadenas b, ya que las a se encuentran en cantidad suficiente a nivel tisular y sanguíneo.
Las glicoproteínas pueden variar en su contenido de carbohidratos. La remoción de residuos de la FSH lleva a la producción de compuestos capaces de unirse al receptor pero no de desencadenar acciones biológicas.
La prolactina consta de 197 a 199 aminoácidos; tiene también variaciones estructurales que incluyen glicosilación, fosforilación y cambios en unión y carga eléctrica. Se encuentran varios tamaños que han llevado a utilizar términos como pequeña, grande y gran-gran prolactina.
Todas estas modificaciones e isoformas llevan a que el inmunoanálisis no siempre pueda reflejar la situación biológica.
están compuestos principalmente por proteínas, pero tienen modificaciones secundarias de carbohidratos y pueden estar selectivamente inmersos en la membrana lipídica, también pueden estar fosforilados, o formar oligómeros por puentes de disulfuro o interacciones covalentes.
Para ejercer su acción, todas las hormonas deben unirse a su receptor específico, estas uniones inician mecanismos intracelulares que conllevan las respuestas celulares. Las hormonas esteroideas y tiroideas son liposolubles y entran a las células libremente y se unen a las proteínas del citosol. Los complejos resultantes translocan al núcleo donde se unen a elementos regulatorios en el DNA estimulando o inhibiendo la transcripción de genes específicos. Todas las demás hormonas se unen a los receptores celulares localizados en la membrana de las células diana. Esta unión disipara uno o más de las vías de transducción que llevan a las respuestas celulares.
Clases y clasificación de Hormonas
Inicialmente las hormonas se clasificaban en tres grupos de acuerdo a su estructura química: hormonas peptídicas y proteicas, las hormonas asteroideas y las hormonas relacionadas con aminoácidos.En vertebrados se clasifican en:
- Aminas
- prostaglandinas
- esteroides
- péptidos y proteinas.
Esteroideas- Solubles en lípidos, se difunden fácilmente hacia dentro de la célula diana. Se une a un receptor dentro de la célula y viaja hacia algún gen el núcleo al que estimula su trascripción.
No esteroideas- Derivadas de aminoácidos. Se adhieren a un receptor en la membrana, en la parte externa de la célula. El receptor tiene en su parte interna de la célula un sitio activo que inicia una cascada de reacciones que inducen cambios en la célula. La hormona actúa como un primer mensajero y los bioquímicos producidos, que inducen los cambios en la célula, son los segundos mensajeros.
No esteroideas- Derivadas de aminoácidos. Se adhieren a un receptor en la membrana, en la parte externa de la célula. El receptor tiene en su parte interna de la célula un sitio activo que inicia una cascada de reacciones que inducen cambios en la célula. La hormona actúa como un primer mensajero y los bioquímicos producidos, que inducen los cambios en la célula, son los segundos mensajeros.
- aminas- aminoácidos modificados. Ej : adrenalina, NE
- péptidos- cadenas cortas de aminoácidos. Ej: OT, ADH
- proteicas- proteínas complejas. Ej: GH, PTH
- glucoproteínas- Ej: FSH, LH

CLASIFICACIÓN
Está hecha a partir de las relaciones anatómicas entre la célula A y la célula B.
1.- Sistémica
La hormona se sintetiza y almacena en células específicas asociadas con una glándula endocrina, esta libera a la hormona al torrente sanguíneo hasta que recibe la señal fisiológica adecuada. La hormona viaja hacia un blanco celular lejano que usualmente tiene una alta afinidad por la hormona. La hormona se acumula en este blanco y se inicia una respuesta biológica que suele resultar en un cambio de concentración de un componente sanguíneo que sirve como señal de retroalimentación para la glándula endocrina que disminuye la biosíntesis y secreción de la hormona. Ejemplo: liberación del hormonas del hipotálamo en un sistema porta cerrado lo que asegura que las hormonas lleguen a la pituitaria anterior, que contiene células receptoras de dichas hormonas.
2.- Paracrina
La distancia entre las células A y B es pequeña de manera que A sintetiza y secreta la hormona que difunde hasta B. Ejemplo: producción de testosterona por las células intersticiales de Leydig, después difunde en los túbulos seminíferos adyacentes.
3.- Autocrina
Es una variación del sistema paracrino en el que la célula que sintetiza y secreta la hormona también es la célula blanco. Ejemplo: prostaglandinas.
4.- Neurotransmisores
Cuando la señal eléctrica de la neurona es sustituido por un mediador químico, (el neurotransmisor) que es secretado por el axón. El neurotransmisor difunde localmente en la sinapsis hasta el receptor de la célula adyacente. Neurotransmisores como acetilcolina y norepinefrina se clasifican como neurohormonas parácrinas.
Está hecha a partir de las relaciones anatómicas entre la célula A y la célula B.
1.- Sistémica
La hormona se sintetiza y almacena en células específicas asociadas con una glándula endocrina, esta libera a la hormona al torrente sanguíneo hasta que recibe la señal fisiológica adecuada. La hormona viaja hacia un blanco celular lejano que usualmente tiene una alta afinidad por la hormona. La hormona se acumula en este blanco y se inicia una respuesta biológica que suele resultar en un cambio de concentración de un componente sanguíneo que sirve como señal de retroalimentación para la glándula endocrina que disminuye la biosíntesis y secreción de la hormona. Ejemplo: liberación del hormonas del hipotálamo en un sistema porta cerrado lo que asegura que las hormonas lleguen a la pituitaria anterior, que contiene células receptoras de dichas hormonas.
2.- Paracrina
La distancia entre las células A y B es pequeña de manera que A sintetiza y secreta la hormona que difunde hasta B. Ejemplo: producción de testosterona por las células intersticiales de Leydig, después difunde en los túbulos seminíferos adyacentes.
3.- Autocrina
Es una variación del sistema paracrino en el que la célula que sintetiza y secreta la hormona también es la célula blanco. Ejemplo: prostaglandinas.
4.- Neurotransmisores
Cuando la señal eléctrica de la neurona es sustituido por un mediador químico, (el neurotransmisor) que es secretado por el axón. El neurotransmisor difunde localmente en la sinapsis hasta el receptor de la célula adyacente. Neurotransmisores como acetilcolina y norepinefrina se clasifican como neurohormonas parácrinas.

INFOGRAFIA

Suscribirse a:
Comentarios (Atom)